


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
皿井 明倫*; 上野 卓哉*; Ngahu, A.*; Ahmad, S.*; 河野 秀俊
no journal, ,
500以上の生物種のゲノム解析から膨大な機能未知の遺伝子がもたらされている。これらのデータから転写因子やそのターゲット予測をゲノムスケールで行うことは、遺伝子発現制御のメカニズムを明らかにするうえで重要である。われわれは、機能未知の遺伝子からDNA結合蛋白質や転写因子のターゲットを予測する方法を開発してきた。DNA結合蛋白質の予測では、DNA結合蛋白質の配列データ,組成やコンテクスト,配列アラインメントの進化的情報や、2次構造,露出表面積,電荷分布の特性(全電荷,双極子等)などの構造情報に基づいて予測する方法を開発した。一方、転写因子のターゲット予測では、蛋白質・DNA複合体の構造データを統計的に解析することにより、アミノ酸と塩基の相互作用による直接認識及びDNAのコンフォメーションによる間接認識の統計ポテンシャルを導出し、それらをもとに予測する方法を開発した。このほか、配列情報を用いる方法,蛋白質・DNA結合データを用いる方法,計算機シミュレーションなどを組合せて精度を上げようとしている。これらの方法をゲノムレベルの予測に応用するため、酵母ゲノムについて解析を行った。この解析のため、まず酵母に関する種々の情報を統合したデータベースを構築した。このデータベースの情報をもとにDNA結合蛋白質や転写因子のターゲット予測を行い、実験との比較解析を進めている。
藤井 聡*; Ara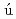 zo-Bravo, M. J.*; 竹中 繁織*; 河野 秀俊; 郷 信広; 皿井 明倫*
zo-Bravo, M. J.*; 竹中 繁織*; 河野 秀俊; 郷 信広; 皿井 明倫*
no journal, ,
タンパク質によるDNA配列の認識には、タンパク質のアミノ酸残基による塩基との直接相互作用による直接認識だけでなく、DNAの配列特異的な構造や物性を通した間接的な認識も重要であることがわかってきた。DNAの構造や物性は塩基配列により異なるため、それらの配列依存性を調べることが重要である。そこで本研究では、DNAの塩基配列ごとに特徴的に存在する構造や柔軟性を調べるため、DNAの水溶液中における動態を分子動力学シミュレーションを実行した。ユニークなテトラマー136種類(AATT, AAAC, CGATなど)を含む12塩基対の二本鎖DNA d(CGCGWXYZCGCG)2 (WXYZ:テトラマー)それぞれについて分子動力学計算を行い、その各10nsのトラジェクトリーからDNAの構造特性を表すステップパラメータを算出し、中央部のユニークなダイマー(AA, AG, GA, GG, AC, AT, GC, CA, CG, TA)ごとにまとめ比較を行った。ステップパラメータの分布に、配列それぞれに特徴的な相関性が観察された。また、ステップパラメータの分布とDNAのリン酸骨格の関係についても報告する。一方、ステップパラメータの分布から平均場ポテンシャルを計算し、そのポテンシャルを用いて、与えられた配列と構造を持ったDNAあるいはタンパク質・DNA複合体中のDNAのコンフォメーションエネルギーと間接認識の特異性の関係解析を行った。
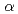 HA1-2mAb由来Fab; 抗原ペプチド複合体のX線結晶解析
HA1-2mAb由来Fab; 抗原ペプチド複合体のX線結晶解析新井 栄揮; 玉田 太郎; 岡村 好子*; 一二三 恵美*; 宇田 泰三*; 黒木 良太
no journal, ,
インフルエンザウイルスの感染は、ウイルス表面の糖蛋白質・ヘマグルチニン(HA)が宿主細胞膜上のガングリオシドやシアル酸含有糖蛋白質受容体へ結合することで始まる。HAは非常に変異しやすく、これがインフルエンザ予防を困難にしている。われわれは、H1型(スペイン風邪,ソ連風邪)やH2型(アジア風邪)ウイルスにおけるHAのアミノ酸配列中に変異しない保存領域を見いだし、その領域に特異的に結合・分解するHA1-2モノクローナル抗体(HA1-2mAb)を開発した。この抗体は抗原を特異的に認識し分解する酵素活性を有しており、「抗体酵素」として新規医薬品・検査診断薬・バイオセンサ開発などへの応用を進めている。われわれはHA1-2mAbの触媒機構を明らかにするために、HA1-2mAb由来Fabと抗原ペプチドの複合体のX線結晶解析を行った。抗体をパパイン消化しFabを調製した後、抗原ペプチド(HAのアミノ酸配列保存領域を含み、18アミノ酸残基から成る)を混合して複合体を調製し、結晶を作製した。X線回折測定では最大分解能2.9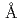 , completeness 96.8%, R-merge 6.6%の回折データを取得した。このデータを用いて分子置換法による構造解析を行ったところ、(1)抗体酵素由来のFabは他のFabとほとんど同じ立体構造をとることや、(2)FabのH鎖とL鎖間のCDR領域付近にペプチドが存在することが確認された。現在触媒基の候補となるアミノ酸残基の同定を実施している。
, completeness 96.8%, R-merge 6.6%の回折データを取得した。このデータを用いて分子置換法による構造解析を行ったところ、(1)抗体酵素由来のFabは他のFabとほとんど同じ立体構造をとることや、(2)FabのH鎖とL鎖間のCDR領域付近にペプチドが存在することが確認された。現在触媒基の候補となるアミノ酸残基の同定を実施している。
玉田 太郎; 木下 誉富*; 黒木 良太; 多田 俊治*
no journal, ,
エラスターゼは、古くから構造情報を基盤とした創薬手法いわゆるStructure-Based Drug Design(SBDD)研究が盛んに行われているセリンプロテアーゼである。したがってエラスターゼの活性部位に存在する触媒基の解離状態を調べることは、SBDDによる創薬に重要な知見を与えると考えられる。そこで、エラスターゼの活性部位の詳細な構造を水素原子の情報を含めて詳細に明らかにするために、超高分解能X線及び中性子構造解析に着手した。結晶化試料はブタ由来のエラスターゼを用い、薬物候補化合物との複合体結晶を作成した。X線回折実験はSPring-8 BL41XUで行い、約0.3mm角の結晶を用いて100Kにて測定した。一方、中性子回折実験は日本原子力研究開発機構の研究用原子炉(JRR3)にある中性子回折計(BIX3)で行い、重水溶液に浸漬させた1.5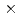 1.30.9mmの結晶を用いて室温で実施した。X線では0.94
1.30.9mmの結晶を用いて室温で実施した。X線では0.94 分解能で、Rmerge=7.5%, completeness=99%、中性子では2.4分解能で、Rmerge=13%, completeness=83%の回折データを収集できた。X線回折データから得られた電子密度では、蛋白質の炭素に結合する水素原子の位置が明瞭に確認できた。また、中性子回折データから得られた核密度では、活性残基であるヒスチジン残基(His-57)のプロトン化の状態が明らかになった。
分解能で、Rmerge=7.5%, completeness=99%、中性子では2.4分解能で、Rmerge=13%, completeness=83%の回折データを収集できた。X線回折データから得られた電子密度では、蛋白質の炭素に結合する水素原子の位置が明瞭に確認できた。また、中性子回折データから得られた核密度では、活性残基であるヒスチジン残基(His-57)のプロトン化の状態が明らかになった。
安達 基泰; 玉田 太郎; 佐藤 勝也; 由良 敬; 鳴海 一成; 黒木 良太
no journal, ,
放射線抵抗性細菌デイノコッカスは、ヒト細胞の約1000倍もの放射線抵抗性を示す。PprAはデイノコッカスより単離された新規なDNA修復促進蛋白質であり、高度放射線抵抗性において最も重要な役割を担っていることが報告されている。本研究は、PprAの構造と機能の関係を解明することを目的としている。大腸菌発現系を用いて調製した組換えPprAを精製し、DNAとの相互作用をアガロースゲルシフトアッセイとゲル濾過により検討した。その結果、PprAは1分子のDNA(pUC19, 2686bp)に少なくとも約280分子の結合が可能であること,PprAとDNAの複合体形成が、低濃度のMg, CaあるいはSrイオンにより促進されること,PprAの多量体構造の形成及びPprAとDNAとの複合体の形成の両方に塩濃度依存性(0-0.4M,酢酸ナトリウム)があることが示された。また、直鎖状の2本鎖DNAとPprAが複合体を形成する場合、PprAの濃度上昇に伴って複合体どうしが会合し、大きな分子量をもつ会合体を形成することが示された。この結果は、放射線によって切断されたゲノムDNAの2つの末端どうしをPprAが近づける役割をもつことを示唆し、PprAの機能発現において重要な知見であると考えられる。
黒木 良太; 本庄 栄二郎; 玉田 太郎
no journal, ,
カイコによる蛋白質発現系は、片倉工業で開発された迅速な発現系である。われわれは目的蛋白質である膜蛋白質の細胞外ドメインの発現を迅速に検討するためにこの系を利用している。そこではまず、目的蛋白質の細胞外領域をアミノ酸配列の相同性と立体構造の予測モデルによって絞り込み、対応するcDNAを、トロンビン切断部位を有するポリアラニンリンカーによって抗体Fc領域に融合したベクターを構築する。このベクターはカイコ発現を実施するバキュロウイルスへのトランスファーベクターであり、カイコへの感染は最終的にはこのトランスファーベクターによって作製された組換えバキュロウイルスにより行われる。カイコの系が迅速なのはこのバキュロウイルスを純化しないで用いることができるからである。これまでに10種類程度の細胞外領域蛋白質の発現を行ったが、ほとんどの蛋白質で発現が確認できた。発現量のカイコによる個体差はあまりなく、安定に発現した。今回のワークショップでは、このうち最も発現量が多かったインターロイキン-13受容体1(IL-13R1)鎖の細胞外ドメインの発現と調製について紹介する。IL-13R1は、アレルギーの発症に関与するサイトカインIL-13の受容体であり、インターロイキン-4受容体(IL-4R)とのヘテロ複合体を構成する。このカイコ発現系によってIL-13R1は、カイコ1頭あたり0.2mg、IL-4Rは、0.01mgの調製が可能であった。本研究は文部科学省タンパク3000プロジェクトにより実施したものである。
由良 敬
no journal, ,
生体中のネットワーク構造として、大きな分子量を持つ定常的な複合体(生体超分子)が重要な位置をしめている。転写装置及びその周辺の分子,mRNAの成熟過程を担うスプライセオゾーム,mRNAからポリペプチドへの翻訳を担うリボゾームなどは巨大分子構造を形成することで機能していることがわかっている。これら生体超分子では電子顕微鏡による構造解析が多くなされている。しかし既存の電子顕微鏡による測定のみでは、一般的には原子分解能の解像度を得ることは難しい。そこで生体超分子を構成するサブユニットの立体構造をX線結晶解析で明らかにし、各サブユニットを電子顕微鏡で得られる全体構造にあてはめることで、生体超分子全体の原子分解能構造を構築することが行われている。この手法による生体超分子の立体構造構築では、サブユニットの構造変化とサブユニット間の相互作用の詳細情報が取り込まれることはまれであり、そのために立体構造の詳細に基づく議論を展開することが難しい。そこでわれわれは、立体構造が詳細に判明している生体高分子複合体において、サブユニット単体のX線結晶解析構造と複合体のX線結晶解析構造を比較することで、複合体形成時にどのような構造変化が起こっているのか、及びどのような部位で相互作用しているかの解析を開始した。抽出された特徴を生体超分子の原子分解能構造構築に取り込むことで、構造構築の精度を向上することを目指している。本講演では、得られた特徴を議論する。
安達 基泰; 玉田 太郎; 佐藤 勝也; 由良 敬; 鳴海 一成; 黒木 良太
no journal, ,
放射線抵抗性細菌デイノコッカスは、ヒト細胞の約1000倍もの放射線抵抗性を示す。PprAはデイノコッカスより単離された新規なDNA修復促進蛋白質であり、高度放射線抵抗性において最も重要な役割を担っていることが報告されている(1)。本研究は、PprAの構造と機能の関係を解明することを目的としている。大腸菌発現系を用いて調製した組換えPprAを精製し、DNAとの相互作用をアガロースゲルシフトアッセイとゲル濾過により検討した。その結果、PprAは1分子のDNA(pUC19,2686bp)に少なくとも約280分子の結合が可能であること,PprAとDNAの複合体形成が、低濃度のMg, CaあるいはSrイオンにより促進されること,PprAの多量体構造の形成及びPprAとDNAとの複合体の形成の両方に塩濃度依存性(0-0.5M,酢酸ナトリウム)があることが示された。また、直鎖状の2本鎖DNAとPprAが複合体を形成する場合、PprAの濃度上昇に伴って複合体どうしが会合し、大きな分子量を持つ会合体を形成することが示された。この結果は、放射線によって切断されたゲノムDNAの2つの末端どうしをPprAが近づける役割を持つことを示唆し、PprAの機能発現において重要な知見であると考えられる。
本庄 栄二郎; 玉田 太郎; 伊倉 貞吉*; 黒木 良太
no journal, ,
創薬の一つの手法として、リード候補化合物溶液に標的タンパク質結晶をソーキングし、そのX線結晶構造を解析することにより、候補化合物の選抜やリード化合物の最適化が行われている。しかしタンパク質結晶は不安定であり、溶媒条件の変化によって損傷を受けやすく、候補化合物が溶解できる条件では構造情報を得ることができないこともある。したがってタンパク質結晶をさまざまな環境で安定化させる技術は創薬の時間やコストの効率化に大きく寄与すると考えられる。われわれはタンパク質結晶を安定化させる手法として分子間ジスルフィド結合に着目し、そのモデルケースとしてT4リゾチーム分子表面に4つのシステイン残基を導入した変異体(S44C/T115C/N68C/A93C、STNA)を作製した。STNAを還元条件下で精製し、さらに結晶化後、徐々に酸化することにより分子間ジスルフィド結合の形成を試みた。その結果、最大で一辺0.4mm程度の結晶が得られ、その回折データを解析することにより、分子間のジスルフィド結合が形成されていることが確認できた。STNAの結晶は純水中でも溶解せず、10% DMSOにソーキングした後でも回折データ収集が可能であった。さらにpH変化にも安定であり、実験を実施したpH4から9の間ですべて1.0から1.5分解能の回折データ収集が可能であった。分子間ジスルフィド結合によって分子間を架橋した結晶は極めて安定であり、さまざまな条件での使用を可能にすることがわかった。